 |
|
|
|
|
|
|
|
|
|
|
Эндогенные
типы покоя семян (Б и В)
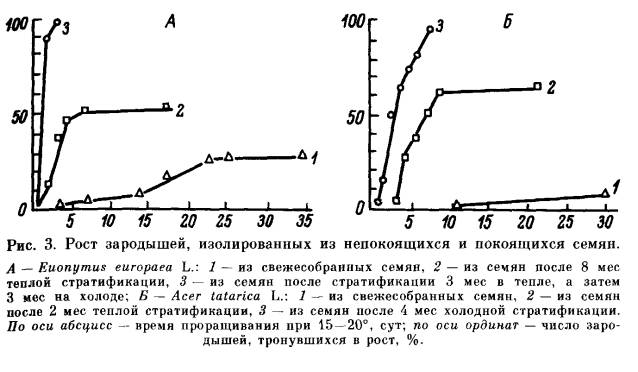
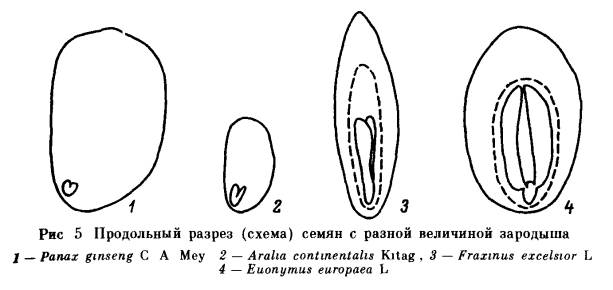
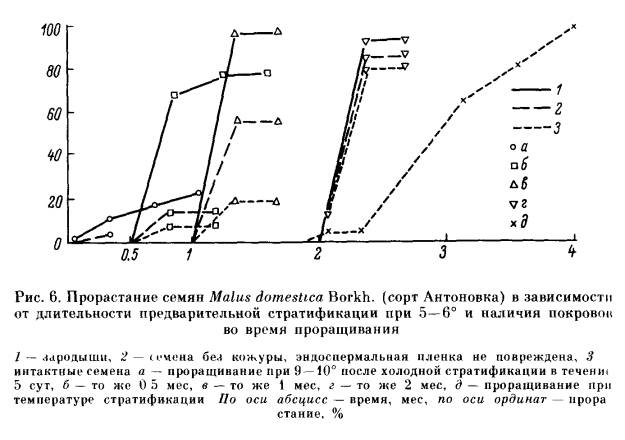
Эндогенный покой семян обусловлен прежде всего свойствами зародыша: его морфологической и анатомической недоразвитостью, или особым физиологическим состоянием, или сочетанием обеих причин. В соответствии с этим выделяют морфологический (Б), разные типы физиологического (В) и морфофизиологического простого (Б -В) и сложного (БВ —В) покоя. Морфологический покой (Б) наблюдается большой частью у растений тропических и субтропических областей, но встречается и у растений нашей флоры, например у некоторых видов аконита (Aconitum). Причиной его является недоразвитость зародыша. Степень этого недоразвития различна у разных растений. Прорастание семян, находящихся в морфологическом покос, может начаться только после того, как в них закончится доразвития зародыша. Этот процесс обычно протекает после отделения семени от материнского растения в условиях теплой стратификации в течение нескольких суток, чаще нескольких месяцев. Физиологический покой (В) широко распространен главным образом у растений, обитающих в умеренном и субтропическом климате. Можно считать установленным, что он вызывается сочетанием особого физиологического состояния зародыша, которое проявляется в пониженной ростовой активности, и недостаточной газопроницаемости тканей, непосредственно его окружающих, в том числе и эндосперма, если он имеется. Такое сочетание создает двойной, или так называемый физиологический механизм торможения. В зависимости от степени торможения роста зародыша и длительности предпосевной подготовки семян мы говорим о разной силе действия ФМТ и, следовательно, о разной глубине покоя: неглубоком (В1), промежуточном (В2) и глубоком (В3) типах физиологического и морфофизиологического покоя. Универсальным фактором, устраняющим ФМТ, является действие пониженной температуры на набухшие семена (холодная стратификация). Во многих случаях, в особенности при неглубоком покое, семена прорастают под влиянием различных других факторов, повышающих ростовую активность зародыша или проницаемость покровов для кислорода и в конечном счете также приводящих к повышению активности зародыша. Неглубокий физиологический покой (В1) характерен для свежесобранных семян огромного числа видов. Он наблюдается у растений, обитающих не только в умеренной и субтропической зонах во и в засушливых тропических областях. Он обычно проявляется в полном отсутствии прорастания или пониженной всхожести семян, а нередко в сужении диапазона температурных или световых условий прорастания. Неглубокий физиологический покой обычно постепенно исчезает в процессе сухого хранения семян, длительность которого варьирует у разных видов от нескольких суток до 5—12 мес. а иногда семена выходят из покоя только после хранения в течение нескольких лет. Наряду с этим известны виды, например лещина (Corylus avellana). у которых семена (без околоплодника) сразу после сбора легко прорастают, но по мере сухого хранения входят в состояние неглубокого покоя. При удалении или хотя бы повреждении покровов зародыши из семян. находящихся в неглубоком физиологическом покос. успешно прорастают и образуют нормальные растения. Обращает на себя внимание многообразие факторов, вызывающих нарушение неглубокого физиологического покоя семян, в большинстве случаев взаимозаменяемых. Одним из наиболее действенных факторов является температурный. Нарушение неглубокого покоя происходит под влиянием кратковременного охлаждения (в течение часов, суток, недель) набухших семян. Сходное действие иногда оказывает сильное прогревание сухих или набухших семян в течение нескольких часов, а также проращивание при переменных температурах. У многих видов семена, характеризующиеся неглубоким покоем, при определенных температурных условиях становятся светочувствительными. Это обстоятельство приходится учитывать при их проращивании. Наиболее типичные светочувствительные семена: сосны обыкновенной (Pinus silvestris), березы (виды Betula), некоторых сортов салата посевного (Lactuca sativa), ряда сорных растений. Реже встречаются растения, у которых свет тормозит прорастание семян: чернушка (Nigella) и др. На сухие семена свет, как правило, не оказывает действия. Исключение представляют семена сосны и некоторых других растений. Длительность облучения зависит от интенсивности и качества cвeтa, видовых особенностей семян и температурных условий проращивания. Акцептором света является содержащийся в семенах пигмент фитохром. О его превращениях под влиянием различных внешних условий можно подробно узнать из книги Быоли и Блэка [224]. Благоприятное действие на прорастание семян, характеризующихся неглубоким покоем, нередко оказывает повышение содержания в атмосфере О2, а иногда СО2. У некоторых видов, наоборот, устранение покоя происходит под влиянием ухудшения условий аэрации или обработки семян дыхательными ядами. Чрезвычайно важно, что неглубокий покой может быть нарушен с помощью различных стимуляторов роста. В частности, на семена некоторых видов положительное действие оказывает обработка растворами нитритов, нитратов (чаще KNO3), тиомочевины, цитокининов, фузикокцина и т. д. Но наиболее постоянное и отчетливое действие вызывают гиббереллины. Значительно сложнее обстоит дело с проращиванием семян, находящихся в промежуточном или глубоком физиологическом покое. К промежуточному типу покоя (В2) относится такой покой, при котором семенам для прорастания необходима довольно длительная (1—3 мес) холодная стратификация. Между тем зародыши, извлеченные из таких семян, обычно растут нормально. Правда, при некотором отклонении от благоприятных условий выращивания у них могут обнаружиться признаки торможения и ненормального роста. Сухое хранение, а также обработка гиббереллином полностью или частично заменяют действие пониженной температуры. Наиболее типичными семенами с таким покоем являются семена абрикоса (Armeniaca), клена ясенелистного (Acer negundo) и ясеня зеленого (Fraxinus viridis). Обработка ГК3 стимулировала прорастание семян у этих видов при условии предварительного удаления околоплодника. Глубокий физиологический покой (В3) обусловлен действием сильного ФМТ прорастания. Если семена с описанными выше типами физиологического покоя имеют зародыши, которые после освобождения от окружающих их покровов большей частью успешно трогаются в рост и обычно дают нормальные растения, то зародыши из семян, характеризующихся глубоким покоем, весьма ограниченно способны к росту. После освобождения от покровов они или не растут совсем, или трогаются в рост медленно (рис. 3), и в большинстве случаев их рост носит ненормальный характер: растут одна или обе семядоли, иногда растет гипокотиль, и на нем возникают опухолевидные образования, но корень большей частью неспособен к росту (рис. 4). Интактные семена, находящиеся в глубоком физиологическом покое, приобретают способность прорастать только после достаточно длительной холодной стратификации. В большинстве случаев ее не удается заменить действием каких-либо других факторов. Правда, теперь обнаруживается все больше видов, у которых удается стимулировать прорастание семян с глубоким покоем с помощью гормональных обработок. Более подробно условия, преодоления сильного ФМТ прорастания рассмотрим после описания глубокого морфофизиологического покоя, в котором этот же механизм оказывает такое же действие и регулируется теми же внешними факторами. Типы простого морфофизиологического покоя (Б — В) характеризуются сочетанием недоразвития зародыша с ФМТ прорастания и, как и типы физиологического покоя, различаются по глубине, определяемой силой действия ФМТ. У большинства изученных видов с морфофизиологическим покоем семян устранению ФМТ, которое происходит в условиях холодной стратификации, должно предшествовать доразвитие зародыша. Степень недоразвитости зародыша у семян, находящихся в морфофизиологическом покое, как и у семян, характеризующихся морфологическим покоем, различна (рис. 5). У лимонника китайского (Schisandra chinensis), женьшеня (Раnах ginseng) и некоторых других растений зародыш зрелых семян находится на самой начальной фазе проэмбрио (т. е. представляет собой недифференцированную группу клеток) или сердечка, а по объему и весу не превышает 0.001 величины семени. У аралии материковой (Aralia continentalis) зародыш достигает фазы Торпедо. В то же время у ясеня обыкновенного (Fraxinus excelsior) и бересклета европейского (Euonymus europaea) зародыш крупный и на вид хорошо дифференцирован. Однако анатомическое изучение показало, что его развитие не завершено полностью [91, 96]. Пока необъяснимым остается тот факт, что длительность периода доразвития зародышей не зависит от того, в какой степени недоразвит зародыш в зрелом семени: у семян женьшеня и бересклета, имеющих зародыш разной степени развития, этот период имеет одинаковую длительность. Процесс доразвития зародыша сопровождается физиологическими изменениями всего семени. В некоторых случаях о них можно судить по так называемому открыванию семян. При этом семя сильно увеличивается в объеме и у него растрескивается кожура или эндокарпий. Открывание семян наблюдается у семян женьшеня, бересклета и многих других растений. В случаях простого морфофизиологического покоя (Б — В) доразвитие зародыша происходит в тепле при 10—25°. Оно может продолжаться от нескольких суток до 3—4 мес, после этого семена должны подвергнуться действию пониженной температуры для устранения ФМТ прорастания Иными словами, нарушение простого морфофизиологического покоя происходит в ходе двухэтап-ной стратификации I этап — в тепле, II этап — при 0 —10°. Часто в естественных условиях за осень в семенах не успевает закончиться процесс доразвития зародыша, следовательно, в зимневесенний период в них не может произойти нарушение покоя Зародыш доразвивается в таких семенах только в течение следующего лета, и, следовательно, торможение прорастания устраняется только во вторую зиму. Важно отметить, что обработка ГК3 семян, находящихся в простом морфофизиологическом покое, стимулирует процесс доразвития зародыша, особенно при неблагоприятных температурных условиях. Так, обработка ГК3 семян видов бересклета позволяет исключить теплый этап стратификации и тем самым сократить общую длительность предпосевной подготовки. Особыми типами простого морфофизиологического покоя являются эпикотильный (Б — Вэ) и двойной покой (Б — Вд) Если У семян, находящихся в физиологическом или обычном морфофизиологическом покое, под контролем ФМТ находятся гипокотиль и корешок зародыша, то при морфофизиологическом эпикотильном покое (Б — Вэ) тормозится рост эдикотиля. Семена с эпикотильным покоем имеет калина обыкновенная (Viburnum opulus). Во время теплого этапа стратификации в семенах происходит доразвитие и появляется корень. Между тем побег трогается в рост только после того, как проростки с достаточно развитым корнем подвергнутся действию пониженной температуры. Простой двойной морфофизиологический покой (Б — Вд) еще более сложен У семян, находящихся в таком покое, например у семян видов Trillium, в покое находятся как гипокотиль, так и эпикотиль зародыша. Для нарушения двойного покоя приходится применять четырехэтапную стратификацию. В течение I этапа в тепле происходит доразвитие зародыша. Затем семена должны подвергнуться во время II этапа действию пониженной температуры для устранения покоя гипокотиля. В течение III (теплого) этапа семена приобретают способность прорастать и образовывать корневую систему. После этого они должны подвергнуться повторному действию холода (IV этап), под влиянием которого устраняется покой эпикотиля и начинается рост побега. Такие семена в естественных условиях прорастают (т. е. образуют нормальное растение) не ранее чем через два года. У семян, находящихся в сложном морфофизиологическом покое (БВ — В), под контролем ФМТ находятся не только прорастание, но и доразвитие зародыша. Устранение действия этого механизма на оба процесса происходит под влиянием пониженной температуры. У одних видов, например видов борщевика (Heracleum), магнолии (Magnolia), стимуляция процесса доразвития зародыша и процесса прорастания происходит наиболее успешно при одинаковой температуре (0 — 3°). Между тем у тюльпана (Tulipa tarda) доразвитие зародыша может происходить при 0 — 10°, но наиболее успешно идет при 9—10°, а прорастают семена, у которых осуществилось доразвитие зародыша, только после стратификации при температуре не выше 3°. Так же как и физиологический покой, типы простого и сложного морфофизиологического покоя могут быть разной глубины в зависимости от силы действия ФМТ. У семян, находящихся в неглубоком морфофизиологическом покое (Б — В1 или БВ — B1), например у семян аралии материковой (Aralia continentalis), нарушение покоя происходит в течение 1—2 мес, а обработка ГК3 не только стимулирует доразвитие зародыша, но и вызывает их прорастание без холодной стратификации. Такое же действие оказывает гиббереллин на семена аралии маньчжурской (A. mandshurica), находящиеся в простом промежуточном морфофизиологическом покое (Б —В2). При простом глубоком морфофизиологическом покое (Б — В3) семена нуждаются в гораздо более длительном пребывании на холоде, а ГК3 хотя и стимулирует доразвитие зародыша, но не заменяет холодной стратификации. В то же время обработка ГК3 не влияет ни на прорастание, ни на доразвитие зародыша семян, характеризующихся сложным глубоким морфофизиологическим покоем (например, у магнолии). Остановимся несколько подробнее на рассмотрении действия сильного ФМТ и условий его преодоления. Стратификационные изменения, приводящие к устранению глубокого покоя, т. е. влияния сильного ФМТ прорастания, обычно протекают при температуре 0 — 7°, не выше, в течение 2—4 мес. У одних видов оптимальными условиями холодной стратификации является более низкая (0—4°), а у других — более высокая (5 — 7°) температура. При несоблюдении благоприятного температурного режима длительность стратификации возрастает, иногда весьма значительно, а энергия прорастания снижается. Так, семена клена татарского (Acer tatarica) при 0 — 3° приобретают способность к прорастанию спустя 3 — 3.5 мес, повышение температуры до 5—7° увеличивает этот срок до 5 мес и более. Стратификационные изменения происходят в покоящихся семенах постепенно: например, у яблони на 15—30-й день пребывания семян на холоде зародыши, освобожденные от покровов, приобретают способность нормально прорастать в тепле; значительно позднее способность к прорастанию появляется у интактных семян (рис. 6). Наиболее благоприятной температурой прорастания стратифицируемых семян является обычно 9—12°, однако чем дольше семена подвергались действию холода, тем более расширяется диапазон температур, при которых они прорастают (табл. 3). Позднее всего семена начинают прорастать в условиях стратификации на холоде. Та6лица 3 Влияние холодной стратификации на расширение
диапазона температур прорастания семян Malus domestica (яблоня домашняя,
сорт Папировка) (по: [52])
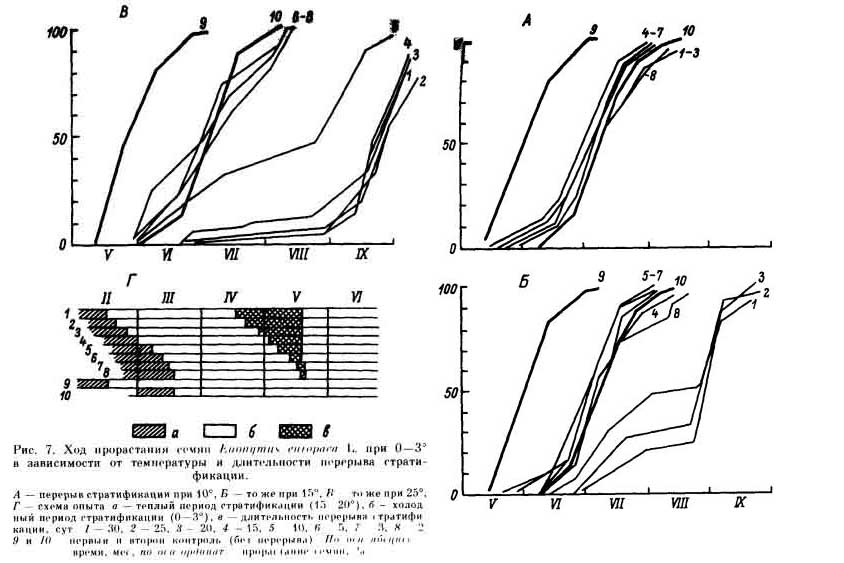
Если семена проращивать в тепле (при 15° и выше) после недостаточно длительного действия холодной стратификации, у семян возникает состояние вторичного покоя. Вторичный покой возникает тем быстрее, чем выше температура, при которой производилось проращивание. Так, у семян клена татарского после 0.5 мес холодной стратификации выдерживание при 15° индуцирует вторичный покой в течение 15 сут, а при 25° — уже в течение 5 сут. Вторичный покой представляет собой по существу реверсию семян к исходному состоянию первичного покоя, а иногда Даже более глубокому, чем первичный покой. Семена, впавшие во вторичный покой, приходится подвергать повторной холодной Стратификации такой же или более длительной, чем для нарушения первичного покоя. Особый интерес представляет температура, близкая к 10°, ее называют компенсационным пунктом покоя — при ней стратификационные изменения у семян с глубоким покоем не происходят, но и вторичный покой не возникает (рис. 7). Ранее упоминалось, что в большинстве случаев холодную стратификацию семян, находящихся в глубоком покое, нельзя заменить действием других факторов. Однако в последние годы появляется все больше и больше данных, свидетельствующих о том, что значительно сократить длительность стратификации или расширить диапазон температур, при которых происходит нарушение покоя, можно с помощью гормональных обработок. В частности, обработка гормонами (кинетином и ГК3) совместно с тиомочевиной (по 2 сут в каждом из трех названных веществ) в 2 — 5 раз сокращает время пребывания семян видов рябин (Sorbus) на холоде, а у многих видов, кроме того, расширяет диапазон температур стратификации с 0 — 3 до 0 — 7° и даже 0—10° [148]. Аналогичные результаты были получены при изучении некоторых видов недотроги (Impatiens parviflora, I. roulei) [149]. У видов клена (Acer) длительную холодную стратификацию можно заменить обработкой семян, освобожденных от околоплодника, раствором кинетина, лучше совместно с тиомочевиной [116, 149]. Если проводить обработку гормонами, растворенными в 20%-ном диметилсульфоксиде, с последующим намачиванием в тиомочевине, то, как показали опыты Л. М. Поздовой, до 70 % семян клена татарского быстро прорастает при температуре 10° и в околоплоднике. Т. В. Далецкая обнаружила, что с помощью обработки ГК3 можно вызвать прорастание в тепле семян некоторых видов купальницы (Trollius), которые находятся в глубоком сложном морфофизиологическом покое и нуждаются в длительной холодной стратификации. Дальнейшие исследования в этом направлении несомненно весьма перспективны. |
||||||||||||||||||||||||||||||||||||||||||||||||
| Информация дана на примитивном агрономическом уровне. Не описаны процессы, происходящие на стадии покоя на физиологическом и биохимическом уровне. Что же происходит с рибосомами, аппаратом трансляции и нуклеиновыми кислотами? Николаева работает все время по этой тематике, ей доступны все новейшие книги и журналы, оборудование и реактивы, не в пример научным сотрудникам из переферии, которые всего этого лишены, а результат жалкое подобие научной работы. Канд. биол. наук А.Ф. Бабицкий Бабицкий А. Ф. ( babandre@mail.ru ) 14.07.2010 13:10 |
| Посещений страницы: 21725 |
|
|
|
|
|
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}